研究概要
哺乳類の雄と雌は形態的にも機能的にも異なっており、雄は精巣で精子を、雌は卵巣で卵子を形成する。精子や卵子の形成には、性染色体(X, Y)上の遺伝子を中心に、雄・雌に特異的に発現する遺伝子群が重要であるが、近年の研究からこれらの遺伝子発現を調節するエピジェネティックな制御が重要であることも分かってきた。我々も個々の遺伝子機能の解析に加えてエピジェネティックな観点から性の分化と配偶子形成の研究を行っている。
Ⅰ 初期胚雌雄判別のシステムを用いた性分化研究
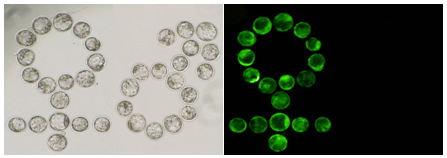
当研究室で作製された142ラインのグリーンマウス(Genomics 2002 )のうち、X染色体上にGFP遺伝子が挿入されたマウス(X-GFPマウス)の雄と、野生型の雌を交配させて得られる初期胚は雌のみが緑色蛍光を発する【図1】。このことを利用すれば普通光では区別できない着床前の胚の段階で雌雄が判定できる(Nat Genet 1998 )。
【図1】X染色体にGFP遺伝子を組み込んだX-GFPマウスを用いた着床前雌雄判定
そこで我々は雌雄を選別した胚を用いて雄雌キメラマウスを作製することで雄と雌になる細胞が入り混じった環境を作り出し、性分化研究を行っている【図2】。通常、受精卵がXY型であれば雄になって精子をつくり、XX型であれば、雌になって卵子をつくるようになるが、雌雄の細胞が入り混じった雌雄キメラでは、精巣と卵巣を併せ持つ雌雄同体もみられたが、ほとんどは雄か雌のどちらかの性で誕生した。
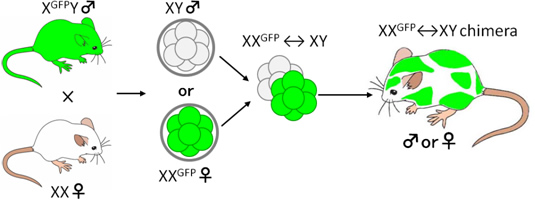
【図2】 X-GFPマウスを用いた着床前の性の判定と雌雄キメラの作製
(この雌雄キメラマウスでは雌(XX型)由来の細胞をGFP陽性細胞として追跡することが可能である)。
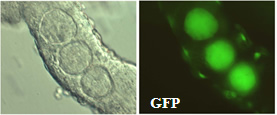
【図2】 雌雄キメラの精細管に存在する卵子様巨大細胞『精巣卵』
雌雄キメラマウスの生殖巣について細胞レベルで詳しく観察すると、精巣中に緑色蛍光をもつ大型の細胞が見られ、雄の精巣中で雌細胞由来の卵子が形成されていることを明らかにした(図3)。さらに、哺乳類において生殖細胞が機能する配偶子へ成熟するために必要なエピジェネティックな現象として知られている、ゲノムインプリントに着目し解析したところ、雌雄キメラの精巣内で見られたXX型生殖細胞のほとんどは、精子型(父型)のインプリントを獲得した精原細胞になっていたが、これらは誕生後消滅してしまうことが分かった。一方で、卵子様に分化した細胞は、不完全ながら卵子型(母型)のインプリントを獲得していることが分かった。すなわち生殖細胞の性分化は、性染色体の組み合わせではなく、生殖細胞の性決定時に運命づけられることを明らかにした (PNAS 2005 )。
Ⅱ 生殖細胞と体細胞の相互作用
生殖細胞は、胎児期にエピブラストの上端部から始原生殖細胞として発生し、未分化な生殖巣原基へと移動したのち、分化した生殖巣(雄なら精巣、雌なら卵巣)の環境に影響を受けて配偶子を形成する。Y染色体は雄特有の染色体であるが、雄への性決定と、生殖細胞において精母細胞への分化に働くことは知られているものの、精子形成における役割は不明な点が多い。我々はY染色体を持たないXX型性転換するマウスを用いて、精子形成におけるY染色体の役割を明らかにしようと研究を行っている。
Ⅲ ノンコーディングRNAと生殖機能
ヒトゲノム上には、およそ22,000のタンパク質をコードする遺伝子が存在するが、それは全ゲノムの2~3%にすぎない。残りの97%の部分は不要な"がらくた"と思われてきたが、近年の研究によりタンパク質をコードしないRNA(ノンコーディングRNA, ncRNA)がゲノム上の至るところから転写されていることが明らかになった。ncRNAの機能については不明なものが多いが、動物個体レベルでどのような生命現象に関わっているのかを我々は生殖現象に焦点を絞って解析を進めている。
miRNAの生殖機能における役割
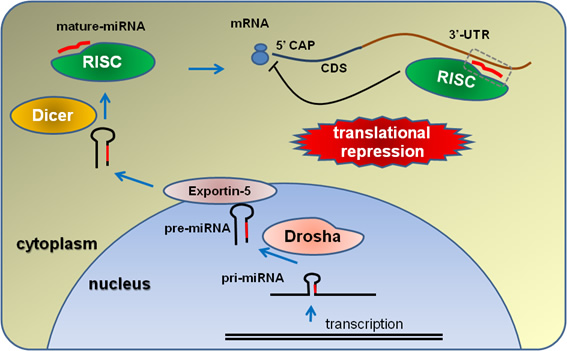
マイクロRNA(miRNA)は20塩基程度の長さをもつノンコーディングRNAで、ヒトでは1048、マウスでは672種類の存在が知られている(miRbase release16)。miRNAは構造遺伝子のイントロンにコードされていたり、ゲノム上のintergenicな領域からpri-miRNAとして転写され、核内でDoroshaやDgcr8により70塩基程度のpre-miRNAとなり細胞質へ移動し、Dicerにより成熟型のmiRNAとなる(図3)。20塩基程度の成熟型となったmiRNAは標的となるmRNAに干渉することで、標的遺伝子のタンパク質への翻訳を抑制する機能を有することが明らかとなっている。
当研究室ではこれまでに報告が無い、生殖機能とmiRNAの関係について、遺伝子改変動物を用い解析を行っている。
【図3】 miRNAの成熟と機能
研究の詳細については各ラボメンバーのHPにも記載されています。